
Une course à relais de 3 neurones
1. Neurone de premier ordre en périphérie : son corps neuronal est dans le ganglion pré-spinal, avant donc d’entrer dans la moelle épinière.
2. Neurone de second ordre ou de transmission vers le thalamus : le corps neuronal est dans la corne postérieure de la moelle et l’axone se projette par les faisceaux spino-réticulo-thalamiques antérolatéraux de la moelle.
3. Troisième neurone : le corps neuronal est dans le thalamus avec des projections au système limbique et au cortex cérébral.
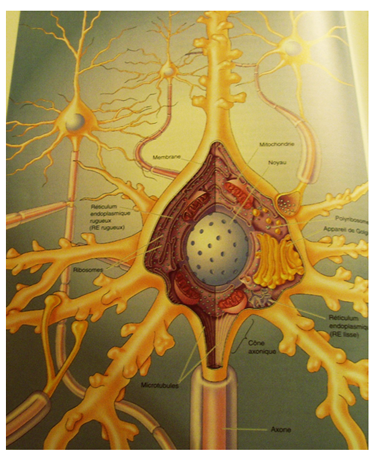
LA GLIE (l’intendance)
Les cellules gliales sont dix fois plus nombreuses que les neurones. Ces cellules remplissent principalement trois fonctions :
1. Elles comblent les espaces situés entre les neurones. Elles constituent donc un tissu de soutien et contribuent à la régulation de la composition du milieu extracellulaire qui baigne les neurones : régulation du pH extracellulaire et de la composition ionique.
2. Certaines cellules gliales sont en contact avec les vaisseaux sanguins. Elles captent dans la circulation sanguine des nutriments nécessaires aux structures nerveuses et assurent ainsi un rôle nourricier.
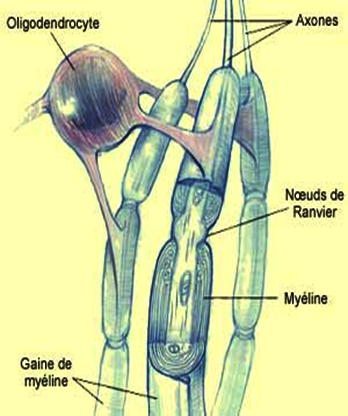
3. D'autres s'enroulent autour des axones, formant des couches de membranes qu'on appelle la gaine de myéline. La gaine de myéline s'interrompt environ tous les deux millimètres, constituant des étranglements que l'on nomme les noeuds de Ranvier. Ce dispositif contribue à rendre plus rapide la propagation des messages nerveux le long de l'axone.
Myélinisation et glie
D'autres serviteurs des neurones, Les Astrocytes
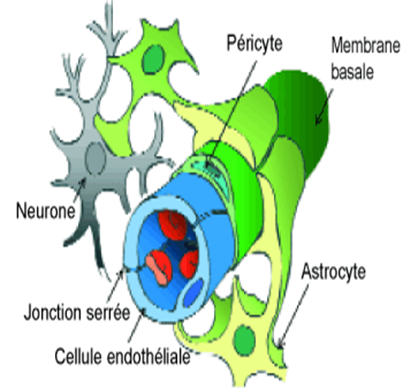
Figure 1. Représentation schématique des composants histologiques de la barrière hémato-encéphalique
Le concept d'une barrière anatomique isolant chimiquement le cerveau de la circulation sanguine s'est progressivement imposé au tournant du XXe siècle, lorsque différents physiologistes (Ehrlich, Biedl, Kraus, Lewandowsky, Goldmann) découvrirent que les colorants vitaux administrés par voie intraveineuse à des animaux d'expérience épargnaient le tissu cérébral, alors que celui-ci fixait les mêmes pigments injectés par voie sous-arachnoïdienne. Par la suite, les travaux de plus en plus précis des neuroanatomistes démontrèrent l'existence d'une série de feuillets séparant rigoureusement le cerveau du flux sanguin, tout au long de l'arborescence vasculaire : le plasma sanguin irrigant le système nerveux central est ainsi confiné à l'intérieur d'un endothélium continu, dépourvu des fenestrations qu'on trouve dans d'autres organes, et dont les cellules sont étroitement soudées entre elles par des jonctions serrées.
Cet endothélium est lui-même enserré entre les prolongements des astrocytes qui s'appliquent tout le long de sa membrane basale, assurant à la fois l'isolation et le soutien tant mécanique que métabolique des neurones, considérés comme cellules «nobles» du tissu cérébral. La construction est encore renforcée par des péricytes pris entre les couches de la membrane basale qu'ils contribuent à sécréter. Le cerveau et la moelle épinière baignent dans le liquide céphalo-rachidien (LCR), dont ils sont toutefois séparés par le feuillet des cellules épendymaires. La barrière hémato-encéphalique se relâche dans certaines régions telles que l'area postrema du quatrième ventricule (centre du vomissement), la neuro-hypophyse ou l'épiphyse.
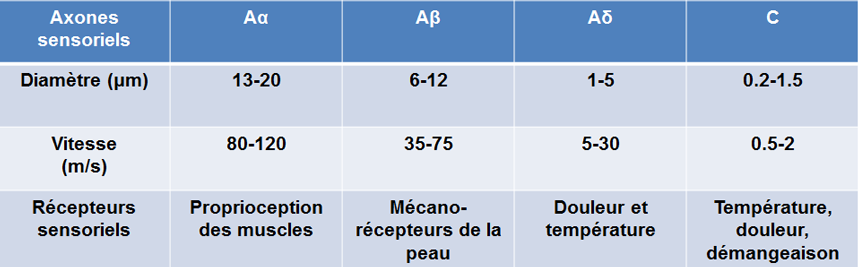
| Les fibres de la sensibilité sont différentes par leur quantité de myéline et leur vitesse de conduction |
|
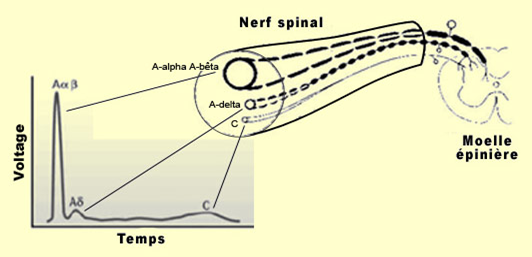
Ce délai provient directement de la vitesse de conduction différente des fibres A-delta et C qui fait que leur message n'atteint pas le cerveau exactement en même temps. Ce qu'on nomme la douleur rapide, qui disparaît assez vite, vient de la transmission d'influx nerveux dans les fibres A-delta (qui conduit l'influx nerveux à la vitesse d'un cycliste). Et la douleur lente, plus persistante, est issue de la stimulation des fibres C non myélinisées
(qui conduit l'influx nerveux à la vitesse d'un marcheur). On estime que ces dernières comptent pour environ 70 % des fibres nociceptives